The digestive system is a group of organs that allows a bird to obtain, digest and absorb nutrients, and remove undigested waste.
The digestive tract includes:
- beak;
- esophagus;
- proventriculus, or glandular stomach;
- muscular stomach;
- intestines (small and thick);
- cloaca;
- liver;
- pancreas.
Beak and oral cavity.
The organs from which the digestive system begins are involved in the collection of food. It is the beak and tongue that are perhaps most characteristic of the parrots. Despite the variety of soft, hard and liquid foods eaten by parrots, all birds of this group have a characteristic large, strongly curved or even hooked beak, which covers a small cup-shaped mandible. The tongue of parrots, usually fleshy, very mobile, plays an active role in manipulating food. The peculiar functions of the beak and tongue distinguish parrots from most other birds, which generally have more delicate and less functional tongues.
The structure of the beak of parrots depends on what they mainly feed on. For example:
- The small compact beak of some small parrots is designed to remove grass seeds from spikelets.
- The relatively large beak of Grays and Amazons is suitable for eating a variety of fruits and seeds.
- The long thin beak of the kea parrot is used for catching insects and pecking meat.
- The massive beak of macaw parrots is adapted for cracking nuts.
Among parrots, lorises and lorikeets seem to have the most specialized beaks. Many species of these birds have a long and thin beak, adapted to obtain pollen and nectar from the calyxes of flowers. Their language is also well adapted to this. It is not thick and fleshy like those of other parrots, but is very long, much thinner, with a “brush” at the end that allows the birds to collect and eat the pollen that makes up most of their diet.
The beak of parrots is also a rather complex instrument. It is represented by a horny sheath (formed by skin from horn, similar to human nails) covering the bony base of the jaws. In most birds, the beak bones are fused with the bones of the skull, but in parrots there is an additional joint between the beak and the skull - the craniofacial hinge. This joint allows for a much greater degree of mobility of the beak.
The horny sheath is constantly built up by skin at the base of the beak and moves forward, replacing those layers of the horn that are erased when hard seeds are chewed.
A complete change of the horny substance of the beak from base to tip occurs in about three months. Various damage to the horny sheath, such as chips or small cracks, gradually move to the tip of the beak and disappear. Deep cracks and damage to the skin at the base of the beak can prevent the growth of a future horn. Therefore, in case of beak injuries, you should immediately consult a doctor. Modern veterinary methods can be quite successful in restoring bird beaks.
In some diseases, the growth of the stratum corneum of the beak is disrupted. Parrot Beak and Feather Disease (PBFD) and various nutritional disorders can cause the beak to flake and crumble. An overgrown beak is usually associated with liver problems.
At the base of the beak, tongue and in the pharynx there are tactile cells, with the help of which birds receive important information about the shape and properties of the surface of the food. Taste buds (though in smaller numbers than in mammals) are located on the root of the tongue. It was possible to establish, mainly in parrots, that they give preference to one or another food not only depending on color and shape, but also on taste.
Goiter.
After food is swallowed, it does not pass directly into the stomach, as in mammals, but first enters a special extension of the esophagus called the “crop” for several hours. Goiter (ingluves) is a sac-like expansion of the esophagus. In most bird species, the crop is located on the right side of the neck and serves to store food and prepare it for digestion. Here, the accumulated food softens, and the carbohydrates it contains are broken down by the enzyme amylase, which is part of saliva. In parrot chicks, the crop is very visible, and the best indicator of the chick's health is the speed at which food passes from the crop to the stomach.
Not all birds have a goiter. In addition to parrots, pigeons, chickens, daytime predators, and sparrows have goiter. Owls, seagulls and penguins do not have goiter.
Birds need a gradual but constant intake of food into the stomach, so the crop allows them to first eat a large amount of food at once, and then ensures that it enters the stomach in small portions.
Stomach.
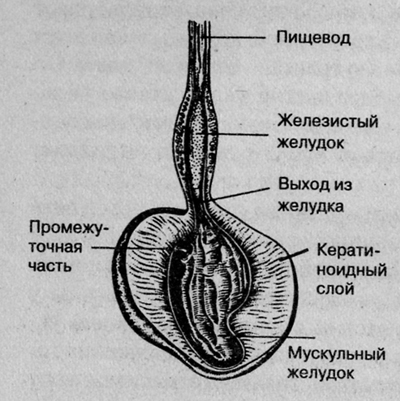
From the crop, food enters the stomach, which in birds is divided into two sections. The first section is the glandular stomach (pars glandularis), called the "forestomach". This thin-walled, spindle-shaped organ is located in front of the muscular stomach. Its walls contain glands that, along with gastric juice, produce digestive enzymes. Between the glandular and muscular stomachs there is an elastic intermediate part - zona intermedia gastris. Here the food mixed with enzymes and acid accumulates before it moves through peristalsis (contraction of the stomach and intestines) into the gizzard. To protect against self-digestion, the zona intermedia gastris is covered with a layer of viscous mucus produced by glands contained in its walls. This mucous layer is important. Because the feed containing acid and enzymes remains here for a long time, the unprotected wall would be damaged. In diseases of the stomach, usually associated with increased acid secretion, the greatest damage occurs in the zona intermedia gastris.
The second part of the stomach - the muscular stomach (pars muscularis) - is also very important. In it, the food is ground due to the movement of powerful muscles of the walls. Small pebbles, which birds swallow from time to time, support the digestion process. Experts debate whether sand is necessary for the gizzard to function properly. Wild birds ingest sand in small quantities. Once in the gizzard, sand remains there for many months and even years, and there is usually no need to regularly replace it. There are known cases when captive birds ate more sand than they should have, especially if they were sick birds, and then their muscular stomach became overloaded. In birds that feed on grain, the muscular stomach is especially well developed. The glands of the muscular stomach produce a secretion that forms a hard keratinoid layer on the stomach wall, which enhances the rubbing effect. In birds that feed exclusively on insects, fruits and nectar, the muscular stomach is poorly developed.
Intestines, liver and pancreas.
The rest of the digestive system—the intestines, liver, and pancreas—is similar in birds and mammals.
From the gizzard, food passes into the intestines, which are divided into two parts: the small and large intestines. In the small intestine, the food lump is mixed with the secretion of the pancreas containing enzymes, as well as with bile secreted by the liver, and moves on. Gut bacteria in the large intestine are essential for breaking down cellulose. Without them, digestion of plant food would be impossible. The food, broken down into its constituent parts, is absorbed through the intestinal wall and is used to provide the body with energy and build its own tissues.
Undigested food remains are excreted through the anus and cloaca in the form of feces (the dark part of the droppings).
The cloaca is a special organ into which the intestines, ducts of the excretory and reproductive systems open. This is where feces come from the digestive tract, urine and white uric acid from the kidneys, and eggs or sperm from the reproductive system. The cloaca consists of three vaguely shaped sac-like compartments that keep various products separate until they are expelled.
Different species of birds have structural features of the digestive system. Thus, some species of parrots lack a cecum and gall bladder.
1. B. Watson, M. Harley “Parrots”, publishing house “World of Books”, 2007.
2. D. Quinten “Diseases of ornamental birds”, Veterinary practice, “Aquarium”, 2011.
The digestive system of birds is divided into the oropharynx, esophagus, stomach, intestines and large wall glands (salivary, liver and pancreas). The general structure of the wall of the digestive tract is the same as that of mammals, i.e. it consists of 4 membranes: mucous, submucosal, muscular and adventitial (or serous). In different departments, the degree of their expression is not the same, some of them may be absent.
The oral cavity does not have lips, cheeks, gums and teeth, and there is no vestibule of the oral cavity. The jaws have been transformed into a beak. The inside of the oral cavity is represented by a two-layer mucous membrane, which has outgrowths and folds. Its multilayered squamous epithelium is keratinized and is located on a thin lamina propria formed by loose connective tissue. The basis of the beak is bone tissue. On the outside it is covered with skin, and on the inside by a two-layer mucous membrane, the epithelium of which in the hard palate is very keratinized. Together with the lamina propria, it forms the palatine papillae. The lamina propria of the mucous membrane of the floor of the oral cavity contains small salivary glands.
The tongue completely occupies the floor of the oral cavity; its root is connected to the hyoid bone. The mucous membrane of the tongue is two-layered: the epithelial layer is formed by stratified squamous, highly keratinizing epithelium; together with the lamina propria on the dorsal surface, it forms numerous filiform papillae. The lamina propria contains complex tubular mucous glands, the excretory ducts of which open on the back, lateral surfaces and root of the tongue, where they are surrounded by taste buds. The muscles of the tongue are transversely striated and are poorly developed in birds (especially chickens).
The oropharynx does not have a velum. Its pharyngeal part consists of 4 membranes: mucous, submucosal, muscular and adventitial. The two-layer mucous membrane forms folds and papillae. Its epithelium is keratinized to a lesser extent than in the oral cavity, and the lamina propria also contains salivary glands and, in addition, diffuse lymphoid formations that form the pharyngeal tonsils. The submucosa also contains the terminal sections of the tubular and alveolar salivary glands, such as the maxillary, palatine, pharyngeal, submandibular, glands of the angle of the mouth and the annular-arytenoid. All of them are predominantly mucous, but mixed and serous terminal sections are found in the maxillary and anterior sublingual glands.
The esophagus is a tube-shaped organ, the wall of which consists of 4 membranes: mucous, submucosal, muscular and adventitial or serous (in cavities).
The mucous membrane is three-layered. The keratinizing stratified squamous epithelium lining it consists of the basal, spinous, granular and horny layers. In the lamina propria, consisting of loose fibrous and a small amount of reticular connective tissue, there are simple tubular branched mucous glands. Their number, degree of branching, and density of arrangement have specific and age characteristics. When the esophagus passes into the glandular stomach, the esophageal tonsil is formed in the lamina propria in the form of a continuous conglomerate of lymphoid tissue, without lymphoid nodules. In the muscular plate, myocytes are located longitudinally, due to which longitudinal folds of the mucous membrane are formed.
The submucosa is very thin. The muscular layer along the entire length of the esophagus consists of two layers of smooth muscle tissue: the internal circular and the external longitudinal. Of these, the inner layer is more developed than the longitudinal one, which is completely absent in ducks. Externally, the esophagus in the cervical part is covered with adventitial membranes, and in the thoracic part - with serous membranes.
Along the esophagus in its cervical region there is a saccular protrusion of the wall - a goiter, in the ventral part of which the goiter groove is located. The wall of the goiter consists of the same membranes as the esophagus, but the epithelial lining in the goiter is thicker. As in the esophagus, there are many mucous glands in the lamina propria. The muscular layer is involved in the formation of sphincters that block the access of food to the crop and glandular stomach. The adventitia and submucosa are without features.
Food is retained in the crop if the stomach is full. Here it softens and is partially subjected to bacterial treatment.
Birds have a two-chamber stomach. The first chamber, where food enters from the esophagus, is called the glandular chamber, and the second is called the muscular chamber. Both chambers differ sharply in morphology and function. If in the glandular chamber food is subjected to enzymatic action with the participation of hydrochloric acid, then in the muscular chamber it is mechanically processed and enzymatic breakdown continues by enzymes that are produced in the glandular chamber.
The wall of the glandular chamber consists of 4 membranes: mucous, submucosal, muscular and serous. The mucous membrane is three-layered. The epithelial layer is represented by prismatic glandular cells, which, plunging into the lamina propria, form the so-called superficial simple tubular glands (or folds, according to some researchers). The glandular epithelium produces a secretion rich in acid glycosaminoglycans, which, covering the entire inner surface of the stomach, protects it from self-digestion. The muscular plate of the mucous membrane is not always continuous.
In the submucosa there are deep glands: monolobular (in ducks) and multilobular (in chickens and geese). The glandular lobules consist of radially arranged tubes lined with single-layer prismatic glandular epithelium with basophilic-stained cytoplasm. The tips of the epithelial cells protrude into the cavity of the tubes, especially when the cells are filled with secretions. The secretion of these cells contains both digestive enzymes and hydrochloric acid. In a word, these cells have double secretion. It is believed that hydrochloric acid is formed in the apical part of glandulocytes, and pepsinogen is formed in the basal part. The tubes open into the central cavity of the glandular lobule - a glandular sac lined with cylindrical cells that produce glycosaminoglycans. From here the secretion enters the cavity of the glandular section of the stomach, where it does not linger and enters the muscular section of the stomach along with food.
The wall of the muscular chamber of the stomach consists of 3 membranes: mucous, muscular and serous. The mucous membrane is three-layered, forms longitudinal and, in the area of the blind sacs, transverse folds. The surface of the mucous membrane is covered with a dense film - cuticle, which is a product of the glandular activity of the muscular part of the stomach. In the lamina propria of the mucous membrane there are simple tubular glands formed by single-layer prismatic glandular epithelium. Its cells produce a complex secretion of protein-carbohydrate nature, which enters the stomach cavity and, mixing with the secretion secreted by the integumentary epithelium of the muscular chamber, solidifies in the form of columns, forming a cuticle with an uneven surface in the form of a grater. Thanks to the presence of this grater and the ingestion of pebbles, mechanical grinding of the food coming here and its chemical breakdown under the influence of gastric juice produced in the glandular chamber occurs.
The muscular plate in the muscular section of the stomach is very thin and, due to the absence of a submucosa, merges with its muscular layer, which consists of 4 muscles: two intermediate and two lateral (ventral and dorsal). The lateral muscles are attached to the tendinous center, where fibrocartilage may be found (in chickens and turkeys). Myocytes contain a large amount of myoglobin, which determines the specific color of the muscles.
The intestine is divided into thin and thick sections. The thin section consists of the duodenum, jejunum and ileum, and the thick section consists of the double cecum and rectum. The rectum opens into the cloaca.
The intestinal wall consists of 4 membranes. The mucous membrane is three-layered. As part of its epithelial layer, there are all types of enterocytes that are found in the intestines of mammals. Together with the lamina propria, the epithelium takes part in the formation of villi and crypts, and due to the presence of the submucosa, folds are formed. The submucosa is very thin, in some places it is almost invisible, and only in the area of the folds is it well expressed. In the loose fibrous connective tissue of the lamina propria and submucosa there is a lot of diffuse and nodular lymphoid tissue.
The structural features of the small intestine include the absence of submucosal intestinal glands in the duodenum, and the large intestine - the presence of both crypts and villi, in addition, the muscular layer in it is represented by a continuous outer longitudinal layer.
The cloaca is an expanded area of the hindgut into which the terminal sections of the digestive, excretory and reproductive systems open. It is divided into an anterior section - the coprodeum, a middle section - the urodeum and a posterior section - the proctodeum. The coprodeum (cavity for feces) is a continuation of the rectum, so the mucous membrane contains villi and crypts. The urodeum is a urinal into which the ureters, the vas deferens in males, or the left oviduct in females open. The surface of the mucous membrane of the urodeum is smooth. The coprodeum and urodeum are lined with single-layer epithelium, while the terminal part of the cloaca, the proctodeum, is lined with stratified skin-type epithelium. The proctodeum contains the copulatory organ (in geese and drakes). In its dorsal part there is a pocket-like invagination - the bursa of Fabricius.
The pancreas of birds is externally covered with a connective tissue capsule and serous membrane, and has a lobular structure. Like mammals, it has external and internal secretion. The exocrine part is formed by tubular, alveolar and alveolar-tubular end sections, which form acini - the structural and functional units of the organ. Their structure is the same as that of mammals. The intrasecretory (islet) part accounts for about 1% of the mass of the gland in birds. A peculiarity of avian pancreatic islets is that some of them consist only of A cells, while others consist only of B cells. In both islets, a third type of morphologically different cells is found, the function of which has not yet been clarified.
The liver of birds (and reptiles) is similar in structure to the liver of lower animals, but, like that of mammals, it has a lobed structure and a similar blood supply. The lobules consist of a network of glandular tubes lined with single-row epithelium (parenchyma). The lumen of the tubes is limited to 5-6 cells. The connective tissue stroma is much less pronounced than in mammals, therefore the liver lobules are approximately determined by the central vein and triads located in the corners of the lobules.
The digestive system in birds has significant structural and physiological features. The poultry digestive system is compact and at the same time extremely efficient. Some small birds process amounts of feed equivalent to approximately 30% of their body weight each day. Birds usually prefer concentrated feed with high energy value, insects and other animals, fruits and seeds. They rarely feed only on plant leaves and grass. For poultry, plant leaves and grass are only an addition to the concentrated diet as a source of vitamins, minerals and some water.
Structural features. Birds lack lips, teeth, and cheeks. The beak-shaped jaws perform the function of capturing food. On the hard palate there are special cone-shaped papillae directed backwards and helping to move food into the esophagus. Similar papillae are found on the tip and back of the tongue; in addition, there are filiform papillae at the root of the tongue. The movements of the tongue and papillae as a whole ensure the advancement of the captured portion of food into the pharynx and entry into the esophagus.
At the bottom and roof of the beak cavity there are small salivary glands. Mixed saliva is a thick and viscous turbid liquid of a slightly alkaline reaction. It contains a lot of mucus, mucin and the enzyme amylase. From 3 to 20 ml of saliva is secreted per day.
Taste sticks in the thickness of the tongue provide the formation of a taste sensation.
Esophagus. Consists of two sections - upper and lower. The upper section starts from the pharynx and ends with the goiter, the lower section starts from the goiter and ends at the glandular stomach. The upper part of the esophagus is longer than the lower part.
Goiter. It is an extension of the esophagus, a cavitary muscular organ before entering the chest cavity. There are sphincters at the entry and exit points. The goiter is well developed in chickens, turkeys, guinea fowl, pigeons; in geese and ducks it is less developed and represents a true expansion of the esophagus.
The esophagus and crop have a well-developed muscular wall. The inner surface of the goiter is lined with stratified squamous epithelium. In the loose connective tissue of its walls there are alveolar-tubular glands that secrete a mucous secretion that does not contain enzymes.
The lower part of the esophagus passes into the stomach.
Stomach. It consists of two sections - the glandular stomach and the muscular stomach.
Its mucous membrane consists of single-layer glandular epithelial cells that produce a special secretion that hardens on its surface, forming a protective layer - the cuticle. The cuticle wears off and is constantly replaced by new secretions. The secret also contains hydrochloric acid.
The food is ground, mixed, and undergoes chemical transformation due to gastric juice and pancreatic juice, bile and intestinal juice periodically thrown out from the intestines. The contents of the muscular stomach enter the duodenum of the well-developed small intestine. The sphincter between them does not close tightly.
The most developed muscular stomach is in geese, which consume a lot of grass while grazing.
Small intestine. It has a spiral shape, characteristic of organs with great functionality. The small intestine is located between the air sacs. The small intestine, like that of mammals, consists of the duodenum, jejunum and ileum. The duodenum looks like a loop. There is no sharp boundary between the jejunum and ileum.
Birds have short intestines compared to mammals. The total length of the thin section in adult chickens is 140... 150 cm, the total length of the entire intestine is more than 170 cm, the digestive tract is 210 cm, i.e. 6 times the length of the body, and in chickens the ratio is 1:4. The total length of the small intestine in ducks is 159 cm, in geese - 234, in turkeys - 205 cm. The length of the duodenum, jejunum and ileum, respectively, in chickens is 30, 102 and 18 cm, in ducks - 38, 116 and 14 cm, in geese -- 46, 165 and 23 cm, turkeys -- 37, 140 and 28 cm.
The intestinal villi are thin, delicate, leaf-shaped and finger-shaped; there are more of them in the duodenum; per 1 cm2 there are 415 of them in chickens, 1512 in ducks, 2051 in geese, and 292 in turkeys. The height of the villi reaches 1 mm, the diameter is 100...200 microns. The surface area of the small intestinal mucosa in chickens is about 2.0 m2, in ducks - 1.5, in geese - 5.5, in turkeys - 8.6 m2.
The pancreatic ducts and bile ducts flow into the duodenum.
Pancreas. It is well developed, has a large size and weight: in chickens 2.8 ± 0.12 g, ducks - 7.9 ± 0.30, geese - 9.1 ± 0.38 g. It has two lobes; in chickens and ducks there are three ducts, in geese and turkeys there are two, which open into one papilla with the bile duct into the duodenum. The pancreas secretes pancreatic juice, which contains the same enzymes as in mammals, but more active.
Liver. Well developed; weight up to 40 g, but can reach up to 1 kg in geese with special fattening. The liver secretes bile that is dark or light green in color; contains alpha-amylase. Bile from the right and left lobes first enters the sinus - expansion of the ducts, then along the sinovesical tract into the gallbladder. The gallbladder and sinus are connected to the duodenum via the vesicointestinal and sinus ducts.
Large intestine. It is represented by caeca and a rectum that opens into the cloaca. The ileum continues into the rectum; At the transition site, two blind processes extend. They are located at an angle of 30° to the intestines and have sphincters. The length of the process is 17...30 cm in chickens, 20...25 cm in geese. Sphincters in the initial and final parts of the rectum prevent the flow of contents from the rectum into the ileum and its premature entry into the cloaca. The rectum is short, poorly developed - in chickens it is 8... 11 cm long, 1... 1.5 cm in diameter. The rectum passes into the cloaca.
Cloaca. It is an expansion of the terminal part of the rectum. The cloaca is divided into three sections by two transverse circular muscles: 1) the anterior, coprodium, or fecal sinus, is a true continuation of the rectum; 2) middle, urodeum, or urinary sinus - the ureters, sperm ducts or oviduct open into it; 3) posterior, proctodium, through which feces and urine are excreted.
In the drake and gander, as well as in the swan, the organ of copulation is located in the cloaca.
The anus in birds has the shape of a slit, surrounded by a ring of obturator muscles.
Physiological features. Feed intake. Birds grab food with their beaks. The food eaten by birds of different species differs in properties. Accordingly, the digestive apparatus of different bird species has its own structural and physiological characteristics.
The captured portion of food is not chewed, but is moistened with saliva and moved into the pharynx by movements of the tongue and further into the esophagus and crop. I
Digestion in the goiter. Characterized by complex motor-secretory function. There are two types of contractions here - peristaltic and tonic. They are difficult to combine and first ensure the flow of food into the left half of the crop, then into the right.
A goiter is characterized by a certain pattern of motor activity: 5...12 consecutive contractions alternating with a pause of 10 minutes. Immediately after filling the crop with food, its movements slow down or stop completely for 35...40 minutes. The movements of the goiter are ensured by contractions of circular and longitudinal smooth muscles; they are regulated by the vagus and sympathetic nerves.
Small components of the goiter contents pass into the lower esophagus in the first minutes, while larger components are retained for up to 14 hours.
The entry of food into the crop is accompanied by stimulation of its glands. In the crop, with the help of its own secretions and saliva, softening and swelling of the feed occurs, as well as the conversion of feed nutrients due to feed enzymes, microorganisms and saliva.
The goiter contains aerobic microorganisms, lactobacilli, E. coli, enterococci, fungi, and yeast cells. Carbohydrates are predominantly hydrolyzed - 8...13% of soluble carbohydrates in the feed, with a small amount of proteins and fats. The end products of carbohydrate transformation are lactic, acetic, propionic and butyric acids.
The main function of the goiter is capacitive.
The movement of contents from the goiter occurs due to small contractions in the area of the goiter funnel. First, one contraction appears, after 1...3 minutes a second wave appears, then 2...3 consecutive contractions, then a long rest.
The bulk of the contents are evacuated from the crop in the first 3...6 hours, a smaller part in the next 8 hours.
Digestion in the stomach. The contents of the crop enter the glandular stomach through the lower section of the esophagus and cause increased secretion of its juice. The secretion of gastric juice occurs continuously. Feed intake stimulates the formation and secretion of gastric juice in chickens up to 11...13 ml/h. Gastric juice contains the enzyme pepsin; There is no lipase in it, since birds do not feed on milk.
The mechanism of stimulation of the gastric glands is neuro-hormonal. The complex-reflex and gastric reflex-hormonal phases of excitation and regulation of the gastric glands have been established. The effect on the gastric glands is realized through the vagus and splanchnic nerves. Protein is a strong stimulant of the gastric glands; Maximum secretion of gastric juice and the enzyme pepsin is observed when the protein content in the diet is within 15...25%. A large amount of protein in chickens, ducks and geese causes overexcitation of the gastric glands and, as a result, inhibition of their secretion.
The glandular stomach also performs a motor function; the rhythm of movement is one contraction per minute. The contents linger here for a short time, no more than an hour, and are mainly saturated with gastric juice. Then it passes into the muscular stomach.
Digestion in the muscular stomach is intense due to the enzymes of the gastric juice of both stomachs and the contractions of the muscular stomach itself. The muscular stomach of birds carries out two types of contractions: phasic and tonic. They appear simultaneously. Against the background of a periodic increase and decrease in muscle tone, a two-phase contraction of the stomach occurs.
The cycle of movement of the muscular stomach begins with the contraction of the superior intermedius muscle. During the period of its shortening, the contraction of the anterior main begins. At the beginning of the relaxation of the latter, successive contractions of the lower intermediate and then the posterior main muscles occur. When the intermedius muscle contracts, the contents of the cranial sac are squeezed into the slit-like cavity between the cuticle plates of the main muscles. Subsequent contractions of the anterior main muscle displace the contents of the slit-like cavity in the posterior direction. Contraction of the inferior intermediate muscle ensures the displacement of the chyme of the caudal sac into the cavity between the main muscles. The posterior main muscle propels the contents towards the cranial cecum.
The main muscles in each cycle of contractions produce counter movements, exerting a rubbing effect on the feed particles. The asymmetry of the arrangement of fibers in the main muscles of the stomach provides the possibility of lateral movements.
At the same time, the chemical transformation of food occurs in the muscular stomach due to the enzymes of juices: gastric, as well as pancreatic, intestinal and bile, which are thrown through a loosely closed sphincter. The sphincter between the muscular stomach and the duodenum periodically opens during digestion, and intestinal contents with enzymes of pancreatic juice, bile, and intestinal juice flow into the stomach. Therefore, proteins, fats and carbohydrates are intensively digested in the muscular stomach. Gastric digestion time is 3 hours.
The inlet and outlet openings in the muscular stomach are located close. In this regard, contractions of the muscular stomach are accompanied by the evacuation of liquid gastric contents, and solid and larger particles of food are retained in the Stomach and undergo deeper transformations. The contents from the stomach enter the intestines in portions and periods.
Digestion in the intestines. Cavity and parietal digestion is carried out with a predominance of parietal digestion. Digestion is characterized by great intensity, since all the enzymes of digestive juices in the intestines are highly active.
The mechanism of excitation and regulation of the secretory activity of the pancreas is reflex-hormonal.
The mechanism of formation and secretion of bile is reflex-hormonal. Hydrochloric acid is a strong pathogen.
Well-developed intestines and villi ensure intensive absorption of converted substances. The total absorption area of chickens reaches on average 2000 cm2. In the intestine, 62...63% of dry substances, 86...91% of protein, 62...54% of fat, 80% of BEV, 30...50% of water are absorbed. Intestinal digestion time 3...5 hours. *
The intestine carries out active contractile activity: the number of peristaltic movements in 15 minutes is 6...10, antiperistaltic contractions are less, from 0 to 3.
Evacuation of the contents occurs in periods of 30...40 minutes, between which there is about 30 minutes of rest. More than 400 ml enters the intestines during the day, and about 250 ml at night. The amount of dry matter in chyme is within 7.5...20%. In the intestinal chyme, high activity of amylase, proteases, and lipases is observed.
The contents in portions - 30...56 portions per hour - enter the blind processes due to the relaxation of their sphincters. The transformation of content substances in the caeca is carried out thanks to enzymes supplied with chyme, its own secretion and the enzymes of microorganisms inhabiting the cacae. The chyme of the blind sacs has amylase and protease activity. In the blind processes, 10...25% of fiber, 8...10% of protein, a small amount of soluble carbohydrates and lipids are broken down.
Digestion in the caecum is accompanied by contractions - 10...12 per 1 hour. In chickens, contractions of the nature of tonic tension are carried out lasting up to 80...100 s.
Periodically, the sphincters open and the contents enter the rectum in portions. For 8...10 contractions of the small intestine, ensuring the flow of contents into the cecum, the latter perform 1 contraction, ensuring the evacuation of the contents into the rectum.
Digestion time in the large intestine is 6...10 hours.
In the rectum, the formation of feces - litter - is completed. The formed droppings are periodically thrown out reflexively through the cloaca.
The most important functions of the intake of substances, their transformation and removal of undigested residues from the body are performed by the digestive system. Birds, being animals adapted for flight, have certain features of their internal structure. They are primarily aimed at reducing their body weight. The digestive system is no exception.
Digestive system of animals
All chordates, which include birds, have a similar structure. It begins. Then, through the pharynx, food enters the esophagus - a tube leading to the stomach. This part of the digestive system is an extension in which food remains for quite a long time. The final absorption of nutrients occurs in the final section of the tract, and the removal of undigested food remains is carried out. It is called the large intestine, which opens outwards into the cloaca or anus.
Features of the digestive system of birds include the presence of a goiter - an expansion of the esophagus and a decrease in the length of the intestine. These structural features contribute to a more intense and reduced body weight of birds.
Can this also be said about other organ systems?
(table).
| Organ system | Structural features |
| Respiratory | Represented by the respiratory tract (nasal cavity, larynx, trachea, bronchi), lungs and air sacs |
| Blood | A four-chambered heart (two atria and two ventricles) and a closed system of blood vessels. Birds have two circles of blood circulation: large and small. Thanks to this, venous and arterial blood does not mix, and birds are warm-blooded animals. Thus, their body temperature does not depend on the ambient temperature. |
| excretory | It consists of paired kidneys and ureters, the ducts of which open into the cloaca. |
| Nervous | The spinal cord and brain, of which the cerebellum is the most developed. |
| Sexual | Birds are dioecious animals. The reproductive system is represented by gonads: testes in males and an unpaired ovary in females, which also serves as a condition for reducing their mass. |
What do birds eat?
The structure of the bird’s digestive system, its features and differences, are primarily determined by the nature of the food. City dwellers feed on insects, seeds and leftover human food.
The taste preferences of birds are quite diverse. Seagulls drink salt water by filtering it through their tonsils.
It is also poisonous due to the characteristics of the food. She eats bugs. Their poison penetrates the skin and feathers of the bird. Although the flycatcher itself has a stable immunity to its action. A person can get burned by touching it. But very small animals die from the harmful effects of flytrap toxins.

Birds that live in natural conditions eat tasty fruits, small vertebrates and even carrion. They have all the equipment for this.
Beak
Birds have no teeth. But they can easily handle both hard nuts and small seeds. After all, it is the beak that initially performs the function of teeth. With its help, birds crush, crush or tear food. The beak consists of a hard horn-like substance that can withstand quite a lot of weight and prolonged loads. For example, a woodpecker can make a hole up to 10 cm deep in the bark of trees to feast on insect larvae.
The shape and size of the beak depend on what the birds eat.

For example, predators have a large curved beak, with which they can easily grab and tear prey. The peregrine falcon even attacks large foxes and wolves. Forest birds extract insects from the bark of trees, making holes in it as if with a chisel.
The tongue, located in the oral cavity, also helps to obtain food. It has special keratinized papillae that help hold food in the mouth.
The bird's digestive system continues with the pharynx, with the help of which partially processed food moves along the tract.
Esophagus and goiter
The bird's esophagus has an unusual structure. It's not just a tube leading into the stomach. It forms an extension called a goiter. It is here that food is retained for a certain time and is chemically processed using saliva. And gulls and pelicans carry food into the crops of the chicks from the feeding site to the nests.

What is bird's milk
The name of this delicious dessert is familiar to everyone since childhood. It got its name due to its external resemblance to a substance that only birds can secrete.
In some birds, a nutritious foamy mass is formed in specialized crop glands. They use this valuable product to feed chicks.
Bird's milk has nothing in common with the milk of animals. It is not liquid at all, but looks like whipped curd mass. But it also contains many valuable organic substances and vitamins that can provide the newborn body with everything it needs.
In penguins, bird's milk is formed in the stomach, and not in the esophagus, and it is richest in fat. This substance is necessary for the development of chicks in difficult northern conditions. And adult penguins use the fat layer not only to warm the body, but also as a supply of food and energy. After incubating the chicks for several months, the males do not leave the laying site. In this state, they are simply not able to obtain food and maintain viability at the expense of subcutaneous reserves.
Features of the structure of the stomach of birds
The stomach of birds also has characteristic features. It consists of two parts.
The sections of the stomach are called glandular and muscular. In the first, food is processed by gastric juice, i.e. Here its chemical processing continues. The surface of the muscular section is covered with a film of keratinized substance. With its help, food is ground to a mushy state.

Surely many have seen birds pecking at small pebbles. They do this not at all because they do not know about their taste. Once in the muscular part of the intestine, stones contribute to the mechanical grinding of rough food. In this way, the lack of teeth in birds is compensated.
Intestines
From the stomach, processed food enters the small intestine, into which the ducts of the liver and pancreas open. In this part of the digestive system, food is finally digested and absorbed into the blood.

The intestine opens to the outside with the cloaca. It is a hole through which undigested food remains, products of the functioning of the reproductive and excretory systems, are removed.
A feature of the intestinal structure is its short length. It depends on the nature of the food. For example, in birds that prefer plant foods, the intestines are 8 times longer than the body. And among insectivorous representatives of birds - only 4.
By the way, the bird that holds the record for the smallest size is considered the most voracious. Did you guess it? Well, of course it's a hummingbird. In a day, she is able to consume pollen, the weight of which is 2 times her own weight.
And the wren brings up to 500 caterpillars and insects to its chicks within one day. What an appetite!
And vultures are the winners among the most greedy and insatiable creatures. With a large amount of food, they fill up so much that it is simply impossible for them to take off. Vultures solve this problem naturally: they wait until their body weight decreases.
Birds digest food intensively, quickly emptying their intestines. This helps reduce their body weight and provide the body with the right amount of energy. An ordinary sparrow can digest a caterpillar in 15 minutes, and grains in 3 hours. This is why birds spend most of their lives searching for food.
Digestive glands
Salivary glands, pancreas and liver - this is the list that birds have. They secrete biologically active substances - enzymes. They speed up the digestion process by breaking down complex carbohydrates into monosaccharides, proteins into amino acids, lipids into glycerol and fatty acids. It is in this form that organic substances can be absorbed by the body.

Features of the digestive process of birds
The internal structure of birds (the table is presented in the article) indicates that all features of their anatomical organization are aimed at adapting to flight. This also applies to the digestive system.
Instead of cheeks and lips, birds have a beak, which is used to capture and hold food. The oral cavity has a hard palate. It is covered by a mucous membrane on which special cone-shaped papillae are located. With their help, food moves into the esophagus. To prevent food from entering the nasal cavity, the palatal fissure is made by the palatine muscles.
Parts of the stomach that perform different functions contribute to intensive food processing. The intestine consists of two sections - thin and thick.
The digestive system of a bird ensures all vital processes of birds. And the peculiarities of its structure determine the ability of these amazing animals to fly.
The digestive system of birds has morphophysiological features associated with adaptation to flight.
Among the physiological features of the digestive apparatus of birds, we note:
- the speed and high intensity of the processes of digestion, absorption and assimilation of nutrients;
- high speed of passage of food masses through the digestive tract. The level of productivity is likely to be higher the higher the rate of feed movement;
- high plasticity and adaptability to the nature of the feed.
The main feed for birds are products of plant (cereal and legume grains, roots, stems and leaves) and animal origin (worms, insects and their larvae).
According to the method of feeding, birds are usually divided into predominantly herbivorous (geese), predominantly carnivorous (ducks) and omnivorous (chickens).
In conditions of intensive industrial production, the traditional division of poultry into groups has, to a certain extent, lost its meaning, since the main feed source is high-calorie feed with a set of basic components of plant and animal origin.
Digestion in the mouth. In chickens, the search for food occurs under the control of vision and touch; olfactory and gustatory perception play a secondary role.
Geese and ducks have a well-developed sense of taste. Geese prefer carrots, carrot tops, and horsetails.
Chickens, turkeys and pigeons have only “daytime vision”, which is due to the absence of “cones” in their retinas. Therefore, the light regime greatly affects the palatability of feed. Chickens, even when hungry, will not peck grain if it is shaded.
One of the characteristics of birds is the absence of teeth. The food is captured by the beak, the shape of which is not the same. In chickens and turkeys it is short, pointed, and hard. Ducks have a soft beak, along the edges there are plates (“teeth”) for straining out food, and on the beak there is a keratinized protrusion (claw or claw) used for tearing off grass. The tongue has keratinized papillae that help to take and hold food.
The number of pecking movements in chickens is 180-240 per minute. In turkeys - 60. In the oral cavity (beak cavity) there are numerous, but poorly developed salivary glands that secrete a little saliva.
By the nature of the secretion, the salivary glands are classified as mucous glands. Saliva contains a lot of mucin, enzymes include ptyalin (amylase) and maltase, but their effect is weak. Due to the fact that food is in the beak cavity for a short time and is not chewed, the effect of amylolytic enzymes in saliva is manifested in the crop. Due to methodological difficulties, the physiology of salivation is poorly developed.
Digestion in the crop. Granivorous birds (chickens, turkeys, guinea fowl and pigeons) have a well-developed goiter - an enlargement of the esophagus. The entrance and exit openings of the goiter are limited by sphincters. In chickens, the crop capacity is 100-120 g of grain, the residence time of the feed in the crop is from 3-4 to 16-18 hours. It depends on the type of feed. Hard and dry food lasts longer than soft and wet food. Liquid food does not stay in the crop.
Ducks and geese have a false crop - an ampulla-shaped expansion of the esophagus.
The mucous membrane of the crop is formed by glandular epithelium; enzymes are not produced.
The crop is a food depot organ; at the same time, the food changes here; it softens, swells, and mixes. Here, partial digestion of nutrients occurs, mainly due to feed enzymes, salivary enzymes and microorganisms supplied with the feed. The main inhabitants of the crop contents are lactobacilli, E. coli, enterococci, fungi, yeast, and ciliates. Microflora breaks down proteins, fats and especially carbohydrates. 15-20% of carbohydrates are digested in the crop. Carbohydrates are fermented quite intensively with the formation of VFAs and lactic acid.
The question of absorption of digestion products by the crop wall remains open. Some authors, taking into account the good blood circulation of the goiter, allow the possibility of absorption of glucose and fermentation products, others believe that absorption does not occur in the goiter.
In pigeons, “goiter milk” is formed in the crop - a white fatty mass, a product of modified epithelial cells of the crop. Pigeons feed their chicks with “goitrous milk” for the first 10-16 days. The milk contains up to 16% protein, 1.3% lipids, mineral salts, vitamin A and B vitamins.
There is a relationship between the filling of the goiter and the stomach. An impulse from an “empty” stomach reflexively causes contraction of the goiter and evacuation of its contents. A “full” stomach inhibits the motility of the goiter. The goiter is innervated by the vagus nerves. Evacuation of the crop contents begins 1-3 hours after feeding. The total duration of food stay in the crop of chickens, turkeys, and pigeons ranges from 3-18 hours.
The main form of goiter contraction is peristaltic contractions. The contractions of the goiter depend on the degree of its filling. An empty crop contracts more often, but with a small amplitude. The motility of the goiter is regulated by sympathetic and parasympathetic nerves. Irritation of the parasympathetic nerves enhances the motility of the goiter, while the sympathetic nerves inhibit it.
Stomach. The stomach of birds is divided into two sections: glandular and muscular. The glandular stomach resembles the simple stomach of mammals and is more developed in birds of prey. In the mucosa there are 30-40 pairs of large tubular glands, the excretory ducts of which open on the folds of the mucosa with special papillae. The volume of the glandular stomach is very small, food is not retained here, and therefore practically no digestion occurs. The glandular stomach is only a “supplier” of gastric juice.
In the gastric mucosa, only one type of secretory cells was found, combining the morphological and functional characteristics of the main and parietal cells. It is assumed that the apical part of the cell produces hydrochloric acid, and the basal part produces pepsinogen. The total acidity of gastric juice in chickens is 0.3%, free - 0.1-0.25%, pH 1.5-2.
In addition to pepsinogen, other proteolytic enzymes were found in gastric juice, in particular gelatinase and gastrixin.
Data on the presence of lipase and especially chymosin in the gastric juice of birds are obviously based on a misunderstanding; the complete absence of milk in the diet of birds makes their presence unlikely.
In 1 hour, 6-9 ml of gastric juice is separated per 1 kg of weight. The secretion of gastric juice is continuous, with wave-like fluctuations in intensity, and intensifies after eating food. The amount of juice depends on the level of food excitability, the type of food (increased when given oats, mixed feed), physiological state (secretion increases during the period of egg production and decreases during molting) and conditions of detention: even a short-term increase in temperature to 35 ° C sharply inhibits juice secretion. Inadequate and monotonous feeding, lack of minerals and vitamins in the diet weaken secretion. Two phases of gastric secretion have been established: complex reflex and neurochemical.
The main processes of gastric digestion occur in the gizzard. This specialized organ is a homologue of the pyloric section of the mammalian stomach, but performs a special function. The muscular stomach is disc-shaped with powerful smooth muscles. Its main function is to squeeze and grind food. Every 20-30 seconds, periodic contractions are observed, as a result of which the food is mixed. The pressure in the stomach cavity reaches 100-160 in chickens, 180 in ducks, and 265-285 mm Hg in geese. Art.
The mucous membrane is keratinized and called the cuticle (keratinoid covering), consisting of a carbohydrate-protein complex similar to mucopolysaccharide. The cuticle has a mechanical significance and protects the stomach wall from the action of pepsin and the penetration of bacteria into the blood. There is no absorption through the cuticle. The cuticle is most developed in birds fed dry and solid food. With prolonged feeding of wet food, the cuticle gradually softens and even disappears.
In the cavity of the muscular stomach there are pebbles, glass, gravel and other foreign bodies - gastroliths, which are used for grinding and grinding food. It is better to give chickens gravel made from quartzite; it is not recommended to replace it with sand, shell, lime, chalk, gypsum, since, when dissolved by hydrochloric acid, they disrupt gastric and intestinal digestion. In the absence of gastroliths, feed digestibility decreases. The main stimulus for contraction during digestion is mechanical irritation of the stomach wall. Regulation of motor activity is carried out by the neurohumoral pathway. The vagus nerve stimulates motor skills.
In addition to grinding food, intense proteolytic processes occur in the muscular stomach. 17-25% of carbohydrates and 9-11% of fats are broken down here. Geese have the highest intensity of gastric digestion; they digest 20-40% of the feed. Gastric emptying in birds occurs reflexively. However, the pyloric reflex of birds is not similar to that of mammals due to the structural features of the sphincter and the presence of an acidic environment on either side of it. In geese, during digestion, chyme enters the intestines continuously, in chickens and ducks - in small portions.
Along with gastric digestion itself, feed is hydrolyzed in the stomach cavity by enzymes that are transported here from the duodenum.
Intestinal digestion. The intestines of chickens are relatively short, divided into thin and thick sections. A feature of intestinal digestion in birds compared to mammals is a higher concentration of hydrogen ions, i.e. lower pH values in all parts of the small intestine.
The basic laws of intestinal digestion and the mechanism for regulating the functions of the main digestive glands are not fundamentally different from those established by I. P. Pavlov’s school for mammals. Therefore, without going into details, we list the main features of intestinal digestion in birds:
- the presence of a powerful enzymatic apparatus of the pancreas,
- intestinal digestion is very intense;
- rapid passage of food through the intestines (in chickens on average 24 hours).
Pancreatic juice is continuously secreted in all types of poultry. Pure juice is a liquid with a specific gravity of 1.0064-1.0108, pH 7.5-8.1. Pancreatic juice of birds has proteolytic, amylolytic and lipolytic activity. Bile in birds is a thick oily liquid of dark green (vesical bile) or bright green (liver bile) color. The amount of bile excreted in birds is higher than in other farm animals except pigs (calculated per kilogram of weight). The process of bile formation in chickens is under neurohumoral control.
The mucous membrane of the small intestine of birds is similar to that of mammals. The structural features of the mucous membrane of birds are the weak development of the submucosal layer and the absence of Brunner's glands in it. In the body of the villi, lymphatic cavities are poorly defined and there are no systems of lymphatic ducts. Intestinal juice in birds contains enterokinase and has amylase, maltase, sucrase and peptidase activity. The vast majority of enzymes in the small intestine have, as in mammals, parietal localization.
The large intestine includes the rectum with paired blind processes. The following processes occur in these processes:
- breakdown of fiber with the participation of microflora enzymes;
- proteolysis processes under the influence of small intestinal enzymes;
- processes of transformation of nitrogenous substances with the participation of microflora;
- synthesis of B vitamins;
- absorption of water and minerals.
Filling of the cecum occurs due to antiperistaltic movements of the rectum and simultaneous peristalsis of the processes themselves. This process occurs periodically, once every 35-70 minutes. The motility of the blind processes is carried out automatically. In the blind appendages of the large intestine, hydrolysis of fiber occurs by cellulolytic microflora, but the ability to digest fiber is limited (10-30% of fiber is broken down). After surgical removal of the cecum, fiber digestion drops to zero.
Colonization of the intestines with microflora occurs after the first intake of food. In addition to cellulolytic bacteria, streptococci, lactobacilli, E. coli and others live in the large intestine. Bacteria hydrolyze proteins, fats and carbohydrates, and also synthesize B vitamins.
It should, however, be taken into account that the microflora of the digestive tract needs the same nutrients as the macroorganism - there is a kind of competition for metabolites. When comparing the effect of feed on the growth of sterile (raised in sterile conditions and receiving sterile feed) and non-sterile (infected with intestinal microflora) chickens, it turned out that sterile chickens grow significantly better than those infected with intestinal microflora.
The large intestine flows into the cloaca, where the openings of the ureters and sperm ducts (or oviducts) also open. The rectum opens into the fecal sinus, where feces are formed. The latter, passing through the urogenital sinus, mixes with urine. Here, uric acid crystallizes and covers the stool with a white coating. In this semi-liquid state, the droppings are released.
Determining the amino acid requirements of poultry. The high egg productivity of chickens, which causes colossal stress on the entire metabolism and primarily protein metabolism, is ensured as a result of the supply of sufficient amounts of protein with feed, which qualitatively corresponds to the needs of the bird. On average, to form one egg and carry out metabolic processes, an adult chicken requires about 10-11 g of digested protein with an amino acid composition similar to the amino acid composition of egg white. It was found that not only a deficiency, but also an excess of amino acids is harmful to chickens.
In an experiment based on feeding chickens with wheat protein poor in lysine, it was shown that when it is added up to 1% relative to the air-dry matter, weight gain and the efficiency of feed nitrogen use increase. If preparations of the latter two amino acids are added to wheat protein, which lacks lysine and contains sufficient methionine and tryptophan, weight gain and nitrogen utilization are reduced compared to the corresponding indicators obtained using the same diet without additives. The introduction of excess lysine into the diet also gives negative results - with an excess of lysine in the feed, chickens show signs of toxicity.
The lack of at least one essential amino acid in the diets of laying hens negatively affects the productivity and quality of eggs. Subsequently, this position was confirmed by many researchers.
Due to the fact that excess and deficiency of amino acids in poultry diets is an undesirable phenomenon, experiments were carried out in which the need of poultry for amino acids, especially essential ones, was established. Balancing chicken diets with respect to essential amino acids and total protein levels allows not only to achieve greater weight gain, but also to increase the efficiency of feed use. When feeding chickens with feed balanced in amino acids, egg production increases and the cost of egg production decreases.
In principle, the methods for determining the amino acid requirements of poultry are the same as for other animals. However, there are some peculiarities in the technique of conducting research on birds. The amino acid requirement is determined mainly in experiments when feeding animals diets with a certain set of feeds (or pure amino acids) deficient in the required amino acid. Diets are formulated with different levels of the amino acid being studied. Results are determined by physiological state, growth, productivity and nitrogen balance. The optimal level of amino acid is taken as the norm of requirement. With an optimal level of amino acids in the diet, the physiological state of the bird is good, its productivity is maximum, young animals give the greatest weight gain, and the highest efficiency of feed and nitrogen use is noted. Dobson, Anderson and Warnick (1964) published interesting studies examining the amino acid requirements of chickens based on simultaneous balancing of all essential amino acids. The essence of their principle boils down to the following. The initial diet is taken with the optimal ratio of amino acids based on existing literature data. Then in the diets the level of all essential amino acids is reduced by 10 or 15% and the level of reduction in weight gain under the influence of each of these amino acids is determined and the average reduction is calculated. If, with a decrease in the content of a given amino acid in the diet, the decrease in weight gain is greater than the average for all groups, then the level of this amino acid in the diet is considered limiting.
As a result of conducting experiments according to the above principle, it was possible to obtain such a ratio of amino acids in the diet of chickens, at which their weight gain was 25% higher than the weight gain obtained on diets compiled on the basis of recommendations for amino acid requirements
US National Scientific Committee on Animal Nutrition (National Research Council, 1962).
Based on amino acid analysis of eggs and muscle tissue, it was concluded that poultry needs for amino acids for growth and egg production are not the same. Requirements for many amino acids calculated from body analysis were similar to those determined in feeding trials. The most significant discrepancies are found in arginine, leucine, tryptophan, methionine with cystine, and phenylalanine with tyrosine. It can be concluded that the method of amino acid analysis of the body gives quite satisfactory results in determining the need for lysine, histidine, isoleucine, threonine, valine and glycine. The method of amino acid analysis of the body with appropriate adjustments for the rate of passage of amino acids through it can be used to determine the bird's need for arginine, leucine, tryptophan, methionine with cystine and phenylalanine with tyrosine.
In the last decade, to calculate amino acid requirements, a method based on the dependence of the content of free amino acids in the blood plasma on the amount of the same amino acids in the diet has been increasingly used. The degree of blood saturation with free amino acids is consistent with their relative concentration in the protein of the feed. The need for amino acids, especially when recalculated per 100 g of crude protein, is not the same in chickens of different directions of productivity. The amino acid requirements of chickens of different productivity levels change with age.
It has been established that the need for essential amino acids increases to a lesser extent than the increase in protein in the diet. It is still difficult to explain the essence of this phenomenon. In addition, experiments have established that the amino acid requirement of chickens and adult birds depends on the energy value of the diet.
Recently, more and more studies have appeared indicating the influence of non-essential amino acids on the need for essential amino acids. It is noted that the inclusion of non-essential amino acids in the diet has a greater effect on weight gain and nitrogen use than the addition of essential amino acids corresponding in the amount of nitrogen. This convinces us of the need to determine the needs of chickens and hens not only for essential amino acids, but also for non-essential ones.
Based on the relationship between the metabolism of arginine and citrulline, arginine and creatine, scientists have established and experimentally confirmed that citrulline and creatine can replace arginine in the diets of chickens and chickens. The inclusion of creatinine and guanidine acetate in the diet also ensures good productivity and increases the content of arginine and creatine in tissues. It also turned out that homocysteine can replace methionine in the diets of chickens and chickens. This is based on its ability to transform into methionine in the presence of methyl group donors. It is also known that methionine in the body of a bird can be converted into cystine. The interconvertibility of cystine and homocysteine has been established. Nicotinic acid in the poultry diet has a sparing effect on the use of tryptophan.
When studying glycine metabolism in chickens, a number of interesting patterns were identified that should be taken into account when determining the need for this amino acid. There is evidence that the inclusion of acetates in the diet of chickens helps to increase the synthesis of glycine in their body. Ethanolamine can also replace glycine in chicken diets. It is assumed that this process occurs through the conversion of ethanolamine into the corresponding glucoaldehyde, which passes into glycolic acid, which is further oxidized into glyoxylic acid. This does not exclude the possibility of direct oxidation of ethanolamine to glycine without intermediate deamination.
It has been established that niacin is directly related to the metabolism of glycine, arginine and alanine. Large doses of glycine in the diet (over 2% of the diet weight) are toxic. When niacin is added, chickens can safely tolerate even 6% glycine in their diets. It is possible that the role of niacin in the metabolism of glycine and arginine is determined by its participation in the metabolism of creatine or the formation of cartilage tissue. Folic acid and cyanocobalamin (vitamin B 12), as well as niacin, mitigate the harmful effects of excess glycine.
If you find an error, please highlight a piece of text and click Ctrl+Enter.